Perspektiven von γδ-T-Zellen für die Krebs-Immuntherapie
DOI: https://doi.org/10.47184/ti.2021.02.051 bis 5 % der T-Zellen im Blut des Menschen exprimieren einen γδ-T-Zellrezeptor anstelle des αβ-T-Zellrezeptors von „konventionellen“ CD4- bzw. CD8-T-Zellen. Im Unterschied zu αβ-T-Zellen erkennen γδ-T-Zellen ihre Liganden unabhängig von HLA-Molekülen. Die im Blut vorherrschende γδ-T-Zell-Population (Vγ9Vδ2-T-Zellen) wird von Pyrophosphaten aktiviert, die von vielen Mikroben sezerniert, aber auch von transformierten (Tumor-)Zellen produziert werden können. Hierdurch werden Tumorzellen im Unterschied zu gesunden Zellen für den Angriff durch γδ-T-Zellen sensibilisiert. Die starke zytotoxische Aktivität verbunden mit der HLA-unabhängigen Erkennung vieler unterschiedlicher Tumoren und der einfachen in vitro Expansion machen γδ-T-Zellen zu attraktiven Effektorzellen für die zelluläre Immuntherapie.
Schlüsselwörter: Adoptiver Zelltransfer, γδ-T-Lymphozyten, Immuntherapie, Phosphoantigene
„Konventionelle“ T-Zellen erkennen über ihren αβ-T-Zellrezeptor prozessierte Antigene in Form von Peptiden im Zusammenhang mit HLA-Klasse-I- (für CD8-T-Zellen) bzw. HLA-Klasse-II-Molekülen (für CD4-T-Zellen). Im Unterschied dazu steht für unkonventionelle Zellen nur eine sehr kleine Anzahl nutzbarer variabler (V) T-Zellrezeptor-Gene im Keimbahn-Repertoire zur Verfügung. Neben unkonventionellen αβ-T-Zellen wie den Mukosa-assoziierten T-Zellen (MAIT) und den Natürlichen Killer-T-Zellen (NKT), die u. a. Lipide erkennen können, sind hier γδ-T-Zellen als wesentliche Gruppe aufzuführen. Etwa 1–5 % der CD3-positiven T-Zellen im peripheren Blut gesunder Erwachsener exprimieren ein γδ-T-Zellrezeptor-Heterodimer. Es gibt lediglich sechs variable Vγ-Gene, die zur Expression eines γδ-T-Zellrezeptors verwendet werden können, und eine ebenso geringe Anzahl potentiell nutzbarer Vδ-Gene. Im Blut dominieren γδ-T-Zellen mit einem Vγ9Vδ2-T-Zellrezeptor, während in epithelialen Geweben einschließlich des Darms vorrangig Vδ1pos- und andere Vδ2negγδ-T-Zellen zu finden sind.
Liganden für den γδ T-Zellrezeptor
γδ-T-Zellen erkennen über ihren T-Zellrezeptor Liganden, die von anderen Immunzellen nicht wahrgenommen werden. Hierzu gehören u. a. Pyrophosphate als Zwischenprodukte der eukaryoten und prokaryoten Cholesterol-Synthese. Solche „Phosphoantigene“ werden von zahlreichen Bakterien und einigen Parasiten (z. B. demMalaria-Erreger Plasmodium falciparum) sezerniert; homologe Moleküle wie das Isopentenyl-Pyrophosphat (IPP) sind auch Zwischenstufen im Mevalonat-Stoffwechsel in eukaryoten Zellen. IPP erfordert jedoch relativ hohe Konzentrationen (im mikromolaren Bereich), um γδ-T-Zellen zu aktivieren. Diese Konzentrationen werden in gesunden Zellen nicht erreicht, können aber von Tumorzellen gebildet werden, in denen häufig eine Störung des Mevalonat- Stoffwechsels vorliegt [1]. Die Überproduktion von IPP gegenüber normalen Zellen hat zur Folge, dass viele unterschiedliche Tumore von γδ-T-Zellen erkannt und abgetötet werden können – woraus sich das große Interesse für denEinsatz von γδ-T-Zellen in der Krebstherapie ableitet. Phosphoantigene werden ausschließlich von Vγ9Vδ2-T-Zellen des Menschen (und einiger Primaten) erkannt; interessanterweise fehlt bei der Maus ein homologer γδ-T-Zellrezeptor. Die selektive Aktivierung von Vγ9Vδ2-T-Zellen durch Phosphoantigene erfolgt zwar unabhängig von HLA-Klasse-I- oder -Klasse-II-Molekülen, erfordert aber zwingend die Expression von Transmembran-Molekülen aus der Gruppe der Butyrophiline (BTN), insbesondere BTN3A1 und BTN2A1 [2]. Die genaue Rolle von BTN-Molekülen wird in dem Beitrag von Th. Hermann in diesem Heft erläutert. Neben Phosphoantigenen sind für Vγ9Vδ2-T-Zellen wenige weitere Stress-assoziierte Liganden beschrieben worden, so z. B. ein ektop auf der Zelloberfläche mancher Tumorzellen exprimiertes DNA-Mismatch-ReparaturproteinMutSH2. Für Gewebe-residente γδ-T-Zellen, die alternativ zu den Blut-γδ-T-Zellen andere Vγ/Vδ-Ketten benutzen (Vδ2neg), sind ebenfalls Stress-induzierbare Liganden auf Tumorzellen identifiziert worden, z. B. Annexin A2 oder der Endothelial-Protein-C-Rezeptor (EPCR) [3]. Ein Überblick zu den bisher gut charakterisierten Liganden für humane γδ-T-Zellen ist in Abb. 1 dargestellt. Wenngleich sich dieser Beitrag auf humane γδ-T-Zellen fokussiert, sei hier erwähnt, dass auch für γδ-T-Zellen in der Maus im wesentlichen Stress-induzierbare Liganden beschrieben worden sind, wodurch sich eine Rolle von γδ-T-Zellen v. a. in der lokalen Immunüberwachung gegenüber infizierten und gestressten Zellen ableiten lässt [4]. Ferner ist von Bedeutung, dass γδ-T-Zellen in aller Regel auch aktivierende NK-Rezeptoren wie NKG2D exprimieren. Da Tumorzellen im Gegensatz zu gesunden Zellen sehr oft entsprechende stressinduzierbare Liganden wie die MHC-Klasse-I-verwandten Moleküle MICA/MICB exprimieren, verfügen γδ-T-Zellen über zwei unabhängige funktionelle Rezeptor-Systeme (T-Zellrezeptor, NK-Rezeptoren), über die Tumorzellen erkannt und eine Effektorfunktion wie z. B. Zytotoxizität ausgelöst werden kann. Interessanterweise kann MICA jedoch auch direkt vom T-Zellrezeptor der Vδ1- γδ-T-Zellen erkannt werden (siehe Abb. 1).
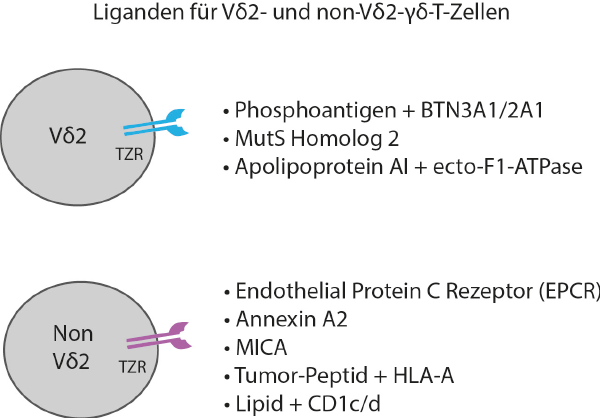
Abb. 1: Liganden für humane Vδ2pos und Vδ2neg γδ-T-Zellen. Für die hier aufgeführten Liganden ist die Erkennung durch den T-Zellrezeptor nachgewiesen. Literaturhinweise hierzu finden sich in den Referenzen 7, 23, 24. Abkürzungen/Anmerkungen: BTN, Butyrophilin; MutS Homolog 2, ektop exprimiertes Homolog des bakteriellen DNA mismatch repair Proteins MutS; MICA, MHC class I-related chain A. Vδ2pos entspricht Vγ9Vδ2, Vδ2neg beinhaltet Vδ1, aber auch andere Vδ2-negative γδ-T-Zellen wie Vδ3. Kürzlich sind auch Beispiele für HLA-A-restringierte Tumor-Peptid-spezifische Vδ1-γδ-T-Zellen beschrieben worden [23].
γδ-T-Zellen in der Tumor-Abwehr
In Mausmodellen der chemischen Karzinogenese wurde beim Vergleich von Wildtyp-Mäusen und γδ-T-Zell-defizienten Mäusen eindeutig die Bedeutung von γδ-T-Zellen für die lokale Immunüberwachung nachgewiesen [5]. Auch für den Menschen leitet sich aus zahlreichen Untersuchungen die Annahme ab, dass γδ-T-Zellen eine wichtige Funktion in der Tumor-Abwehr haben. So können γδ-T-Zellen wie bereits erwähnt unterschiedliche Tumorzellen erkennen und abtöten, so z. B. Leukämien und Lymphome, aber auch solide Tumore unterschiedlichen Ursprungs [6]. Die Erkennung von Phosphoantigen-produzierenden Tumorzellen durch Vγ9Vδ2-γδ-T-Zellen ist abhängig von BTN3A1/2A1-Molekülen, was durch den Einfluss von entsprechenden antagonistischen anti-BTN-Antikörpern gezeigt werden konnte. Aktivierte Vδ1-γδ-T-Zellen, die ebenfalls starkes zytotoxisches Potential gegenüber Tumorzellen haben, werden dagegen vorrangig über die hoch exprimierten aktivierenden NK-Rezeptoren wie NKp30/NKp44 aktiviert („DOT-Zellen“) [7].
Neben der HLA-unabhängigen Tumorzell-Erkennung spricht auch die regelmäßige Präsenz von γδ-T-Zellen innerhalb tumorinfiltrierender Lymphozyten (TIL) für eine Rolle von γδ-T-Zellen in der Auseinandersetzung zwischen Immunsystem, Tumor und Tumor-Mi-croenvironment. Als starkes Argument für eine wichtige protektive Rolle von γδ-T-Zellen in der Tumorabwehr wurde in einer umfassenden Studie die Korrelation zwischen tumorassoziierter γδ-T-Zell-Häufigkeit und vorteilhafter Prognose bei 39 unterschiedlichen Tumor-Entitäten gewertet [8]. Es handelte sich hier um eine bioinformatische Analyse von Transkriptomen aus 18.000 Tumor-Proben, basierend auf dem CIBERSORT Algorithmus. Die rechnerische Abundanz von γδ-T-Zell-Transkripten war in dieser Studie jedoch ungenau. Demzufolge musste die Einschätzung, dass tumor-assoziierte γδ-T-Zellen im Vergleich zu insgesamt 22 unterschiedlichen Immunzell-Populationen bei einer Vielzahl von Tumoren der der prognostisch günstigste Faktor sind, etwas relativiert werden. Dies ist in nachfolgenden Studien erfolgt, wobei aber dennoch eine positive und statistisch signifikante Korrelation zwischen der Anzahl tumorinfiltrierender γδ-T-Zellen und dem Überleben bei bestimmten Tumoren wie Colon- und Prostata-Karzinom bestehen bleibt [9].
γδ-T-Zellen verfügen über eine erstaunliche funktionelle Plastizität. Während frisch aus dem Blut isolierte γδ-T-Zellen in der Gewebekultur nach Aktivierung i. d. R. Zytokine wie Interferon-γ, Tumor-Nekrose-Faktor-α und zytotoxische Mediatoren wie Granzyme produzieren, können sie unter entsprechenden polarisierenden Bedingungen – ähnlich wie αβ-T-Zellen – auch andere Botenstoffe wie IL-4 und IL-9 sezernieren und suppressive Aktivität erwerben. Die Differenzierung von T-Zellen einschließlich γδ-T-Zellen wird auch durch epigenetische Prozesse gesteuert [10]. Neben seiner Eigenschaft als Anti-Oxidant beeinflusst z. B. Vitamin C die Differenzierung von γδ-T-Zellen über die Aktivierung der sog. „ten-eleven translocation“(TET)-Enzyme. Vitamin C erhöht die Effektorfunktion von γδ-T-Zellen [11], stabilisiert aber auch in Gegenwart von Transforming Growth Factor β die Expression und Demethylierung von FOXP3 und induziert dadurch regulatorische Aktivität in γδ-T-Zellen [12]. Im Zusammenhang mit der Tumorabwehr ist ferner von Interesse, dass aktivierte Vγ9Vδ2-T-Zellen nach Tumorzell-Abtötung Fragmente von absterbenden Tumorzellen phagozytieren und nach Prozessierung entsprechende Peptide im Sinne des „cross-priming“ HLA Klasse I-restringierten tumorspezifischen CD8 αβ-T-Zellen präsentieren können [13]. Somit könnten γδ-T-Zellen nicht nur für die direkte Tumorzell-Eliminierung von Bedeutung sein, sondern auch als Initiatoren einer nachgeschalteten Rekrutierung von „konventionellen“ tumorspezifischen αβ-T-Zellen.
Die Plastizität von γδ-T-Zellen könnte prinzipiell allerdings auch negative Auswirkungen für die Tumorabwehr haben. So ist z. B. beschrieben worden, dass Vδ1-γδ-T-Zellen innerhalb der TIL bei Brustkrebs die tumorreaktiven αβ-T-Zellen supprimieren können [14]. In einer anderen Studie sind allerdings genau solche Vδ1-TIL als prognostisch günstiger Marker bei Frauen mit „triple-negativem“ Brustkrebs identifiziert worden [15]. Aus Maus-Modellen ist darüber hinaus bekannt, dass IL-17 produzierende γδ-T-Zellen einen pro-tumoralen Einfluss ausüben können, was u. a. insbesondere auf die Rekrutierung von neutrophilen Granulozyten durch IL-17 zurückgeführt wurde. IL-17 ist offensichtlich ein Zytokin, das die Tumor-Propagierung begünstigen kann [7]. Gerade in Bezug auf IL-17-produzierende γδ-T-Zellen gibt es aber bedeutsame Unterschiede zwischen Maus und Mensch: γδ-T-Zellen gehören beim Menschen (im Gegensatz z. B. zu intestinalen γδ-T-Zellen in der Maus) nicht zu den vorrangigen IL-17-Produzenten. Insgesamt wird aber die Aktivität von lokalen γδ-T-Zellen – wie auch von anderen Immunzellen – durch das im allgemeinen immunsuppressive Tumor-Mikromilieu beeinträchtigt. Zahlreiche Faktoren können hierzu beitragen, so z. B. immunsuppressive Zytokine wie TGF-β, inhibitorische Stoffwechselprodukte wie Prostaglandin E2 und Kynurenine, inhibitorische Lektine wie Galektine, oder auch die Expression von PD-L1 auf Tumor- oder Stromazellen [16].
Strategien zum klinischen Einsatz von γδ-T-Zellen
Prinzipiell werden drei Strategien zum klinischen Einsatz von γδ-T-Zellen verfolgt, nämlich erstens der adoptive Transfer von in vitro expandierten γδ-T-Zellen, zweitens die in vitro Aktivierung von γδ-T-Zellen, im Wesentlichen über Zoledronat, monoklonale Antikörper oder bispezifische Antikörper-Konstrukte, sowie drittens die Entwicklung von genmodifizierten Effektorzellen (CAR (Chimeric Antigen-Receptor)-γδ-T-Zellen, TEG (αβ-T-Zellen mit Expression eines definierten γδ-T-Zellrezeptors) (Abb. 2).

Abb. 2: Aktivierung von tumorreaktiven Vδ2pos und Vδ1pos γδ-T-Zellen. Links: Vδ2pos (= Vγ9Vδ2) T-Zellen können selektiv durch Aminobisphosphonate wie Zoledronat, durch synthetische Phosphoantigene sowie durch agonistische anti-BTN3A1-Antikörper aktiviert werden. Rechts: Die Aktivierung und Expansion von Vδ1pos T-Zellen („DOT“) erfordert optimierte Protokolle mit initialer anti-CD3-Antikörper-Aktivierung und anschließender sequentieller Gabe von Zytokinen wie IL-4, IL-15, IL-1β und Interferon-γ. Sowohl Vδ2pos als auch Vδ1pos γδ-T-Zellen exprimieren aktivierende NK-Rezeptoren, Vδ2pos vorranging NKG2D, Vδ1pos zusätzlich NKp30 und NKp44. Unten: Die Aktivierung und Effektorfunktion von γδ-T-Zellen wird maßgeblich beeinflusst durch bispezifische Antikörper, epigenetische Modulatoren wie Vitamin C, Decitabine, Histondeacetylase (HDAC)-Inhibitoren, darüber hinaus durch Modulation des Tumor-Mikromilieus und ggf. durch Checkpoint-Inhibitoren. Ferner befinden sich genmodifizierte T-Zellen wie CAR-γδ-T-Zellen und TEG-Zellen (αβ-T-Zellen mit hochaffinem γδ-T-Zellrezeptor) in der Entwicklung.
Die in vitro Aktivierung und anschließende in vitro Expansion lässt sich auch unter GMP-Bedingungen durchführen. Eine einfache aber hochselektive Methode zur Aktivierung (nach Zusatz von Wachstumsfaktoren wie IL-2 und/oder IL-15) von Vγ9Vδ2-T-Zellen ist die Verwendung von Aminobisphosphonaten wie Zoledronat (Zometa©), die – neben ihrer Wirkung auf die Knochenresorption – auch den Mevalonat-Stoffwechselweg beeinflussen und hier zu einer Akkumulation des γδ-T-Zellrezeptor aktivierenden Phosphoantigens IPP führen. Ausgehend von Ficoll-Hypaque-separierten mononukleären Zellen des peripheren Blutes (PBMC), die etwa 1–5 % γδ-T-Zellen enthalten, expandieren γδ-T-Zellen nach Zoledronat-Aktivierung und IL-2-Zusatz zu > 90 % innerhalb von 2 Wochen [17]. Neben Aminobisphosphonaten werden auch hochpotente synthetische Phosphoantigene für die in vitro Expansion von Vγ9Vδ2-T-Zellen eingesetzt. Die Expansion von Vδ1-γδ-T-Zellen ist aufwändiger und erfordert neben der T-Zellrezeptor-Aktivierung über anti-CD3-Antikörper den selektiven und sequentiellen Zusatz mehrerer Zytokine einschließlich IL-1β, IL-2, IL-4, IL-15, Interferon-γ. Die so generierten DOT(„Delta One T cell“)-Zellen sind aufgrund ihrer starken Expression aktivierender NK-Rezeptoren zytotoxisch gegenüber Tumorzellen einschließlich der chronischen lymphatischen Leukämie (CLL) [7].
Alternativ zu dem adoptiven γδ-T-Zell-Transfer sind Strategien zur in vitro Aktivierung von γδ-T-Zellen in der klinischen Entwicklung. Aminobisphosphonate sind zugelassene Medikamente zur Behandlung von Knochenerkrankungen wie Osteoporose bzw. Knochenmetastasen von Tumoren. Zusammen mit niedrig dosiertem IL-2 kann Zoledronat zu einer nachweisbaren in vitro Aktivierung von γδ-T-Zellen führen, was in kleineren Studien auch mit einem positiven Effekt auf die Tumorerkrankung (z. B. beim Prostata-Karzinom) verbunden war [18]. Eine direktere Aktivierung von Vγ9Vδ2-T-Zellen als mit Zoledronat wird durch agonistische anti-BTN3A1-Antikörper erzielt [19]. Ein agonistischer humanisierter anti-BTN3A1-Antikörper befindet sich z. Zt. in der klinischen Phase-I-Studie, mit der potentiellen Indikation der Aktivierung und Mobilisierung von γδ-T-Zellen bei Patienten mit soliden Tumoren. Ferner befinden sich bispezifische Antikörper in der Entwicklung, die – basierend auf dem erfolgreichen Konzept der CD19xCD3 „bispecific T-cell engagers“ (BiTE) – variable Vγ- oder Vδ-Ketten mit Tumorantigenen vernetzen und dadurch die Lyse von Tumorzellen durch entsprechende γδ-T-Zellen auslösen [20]. Neben der Identifizierung von relevanten (und möglichst selektiven) Tumorantigenen wird hier vor allem auch an der Optimierung der Antikörperkonstrukte („nanobodies“) gearbeitet. Ferner kann die Effektorfunktion von γδ-T-Zellen auch durch Fab-Fragmente von bestimmten anti-CD3-Antikörpern wie UCHT1 selektiv verstärkt werden [21]. Schließlich werden auch Konzepte zur gentechnischen Veränderung von T-Zellen verfolgt. So befindet sich der Einsatz von γδ-T-Zellen als Rezipienten für chimäre Antigen-Rezeptoren (CAR) in Erprobung. CAR-γδ-T-Zellen haben gegenüber „konventionellen“ (d. h. αβ-T-Zellen) CAR-Zellen den theoretischen Vorteil, dass außer dem exprimierten CAR auch der endogene Vγ9Vδ2-T-Zellrezeptor zur Tumorzell-Erkennung beitragen kann [22]. Eine weitere vielversprechende Strategie besteht darin, γδ-T-Zellrezeptoren mit hoher Affinität für Tumorzellen zu selektionieren und diese in αβ-T-Zellen zu transduzieren. Dieses Prinzip der TEG („αβ-T-cells with expression of defined γδ-T-cell receptor“) befindet sich im frühen Stadium der klinischen Erprobung [23]. Eine Übersicht zu möglichen Strategien für den klinischen Einsatz von γδ-T-Zellen ist in Abb. 2 dargestellt.
Chancen und Herausforderungen
Es gibt gute Gründe für die derzeitige Begeisterung, γδ-T-Zellen in die klinische Anwendung zu bringen – was sich u. a. auch ablesen lässt an der zunehmenden Anzahl von Firmen, die sich auf γδ-T-Zellen fokussiert haben. Aktuelle Übersichten hierzu finden sich in den Referenzen 23 und 24, Übersichten zu klinischen Studien in Referenzen 6 und 23. Ein überragender Vorteil von γδ-T-Zellen ist die aufgrund der HLA-Unabhängigkeit offensichtlich gefahrlose Anwendung von allogenen γδ-T-Zellen für den adoptiven Zelltransfer. In einer kürzlich durchgeführten Phase-I-Studie erhielten 132 Patienten mit unterschiedlichen soliden Tumoren insgesamt 414 Infusionen von allogenen (d. h. von gesunden Probanden etablierten) Vγ9Vδ2-T-Zellen, ohne dass schwerwiegende Zwischenfälle wie graft-versus-host disease beobachtet wurden [25]. Somit eröffnet sich die Perspektive, γδ-T-Zellen als „off the shelf“-Zellbank-Produkt zum Einsatz bei Tumorpatienten herzustellen. Fraglich ist jedoch, ob sowohl der adoptive Zelltransfer als auch die Applikation von γδ-T-Zell-spezifischen bispezifischen Antikörpern jeweils für sich allein genommen therapeutisch ausreichend effektiv sein werden. Die Kombination beider Strategien liegt nahe. So könnten z. B. in vitro expandierte γδ-T-Zellen vor dem adoptiven Transfer mit bispezifischen Antikörpern „beladen“ werden, um die transferierten γδ-T-Zellen noch effizienter Tumorzellen angreifen zu lassen [24].
Ein wichtiger weiterer (wenn auch nicht γδ-T-Zell spezifischer) Aspekt ist die Frage, wie das suppressive Tumor-Mikromillieu beeinflusst werden kann, um die Wirksamkeit von γδ-T-Zell-Therapien zu erhöhen. Strategien zur Blockierung inhibitorischer Moleküle wie Galektine sind hier ebenso zu betrachten wie auch die (lokale) Applikation von Adjuvantien und Immunstimulanzien wie Agonisten für Toll-like- und STING-Rezeptoren [24].
Autor
